Per vari decenni i ricercatori hanno studiato i meccanismi molecolari che regolano i ritmi circadiani, le oscillazioni giornaliere reputate ormai ubiquitarie in biologia.
Questo orologio di base è stato ben analizzato nel contesto delle cellule animali. Si tratta prevalentemente di proteine capaci di generare un feedback a loop nel processo trascrizione/traduzione che pertanto guida le oscillazioni circadiane dell’espressione genica e dei pathway biosintetici cellulari.
Gerarchia del ritmo circadiano
Tutti questi orologi cellulari presenti nei tessuti periferici sono gerarchicamente sincronizzati da un master clock presente nel sistema nervoso centrale e rappresentato dal nucleo soprachiasmatico situato a livello ipotalamico. Esso permette di generare delle risposte in base al quantitativo di luce e a stimoli primitivi quali la fame. Nuovi studi, comunque, rivelano che in modelli murini l’assenza di un importante gene circadiano, Bmal1, permette comunque di generare dei ritmi giornalieri endogeni, dimostrando quindi che potrebbero esistere degli altri master clock nel corpo che regolano l’espressione genica. In letteratura scientifica esistono numerose evidenze del ritmo circadiano, anche se non tutti i meccanismi classici descritti sono in grado di spiegare tutti i fenomeni. Nei cianobatteri c’è un loop post-trascrizionale basato sulla fosforilazione dei substrati che guida i processi circadiani di trascrizione e traduzione. Nelle piante e nei funghi, esistono tali loop e sono simili a quelli presenti negli uomini. Inoltre, in tutti i vari regni cellulari, esistono delle peculiari oscillazioni circadiane relative all’ossidazione e alla riduzione delle perossiredoxine, una famiglia di enzimi antiossidanti, che proseguono il loro ritmo circadiano anche in assenza di trascrizione e traduzione.
Nelle cellule mammifere Bmal1 codifica per un attivatore trascrizionale che è essenziale per il loop a feedback del ritmo tradizionale. Esperimenti passati avevano dimostrato che prelevando dei campioni di pelle o di fegato privi di BMAL1 e coltivandoli in vivo con desametasone (glucocorticoide sintetico noto per modulare i ritmi circadiani), robusti ritmi nell’espressione di migliaia di trascritti e di centinaia di proteine continuavano per giorni dall’inizio del trattamento.

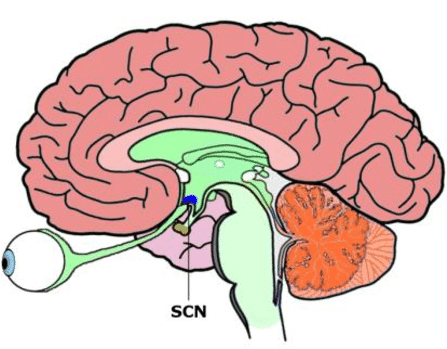
Anatomia del nucleo soprachiasmatico ipotalamico
Il piccolo nucleo soprachiasmatico (SCN) dell’ipotalamo gioca un ruolo centrale nella programmazione giornaliera delle funzioni dell’organismo, regolando di giorno in giorno le oscillazioni delle attività cellulari e sincronizzando le stesse in base ai cicli di luce e di buio. Questo orologio biologico guida l’espressione giornaliera di funzioni omeostatiche vitali anche diverse fra loro, come nutrirsi, bere e termoregolarsi. L’SCN impone un ordine temporale mediante la genesi di segnali di output che forniscono informazioni relative all’orario della giornata e attraverso l’accumulo di informazioni in entrata in grado di mettere a punto questo orologio intrinseco del corpo.
I segnali luminosi provengono dalla retina e sono trasmessi al nucleo ipotalamico mediante i tratti retino-ipotalamici. I neuroni del SCN contengono differenti neurotrasmettitori quali somatostatina, VIP, ADH e neurotensina. I pathway efferenti, incluso uno che regola la funzione della ghiandola pineale secernente melatonina, regolano vari ritmi circadiani comportamentali, endocrini, cardiovascolari e termoregolatori.
Da un punto di vista anatomico il SCN è una struttura neuronale ovviamente pari posizionata nell’ipotalamo anteroventrale, su ciascuno lato del terzo ventricolo, appena superiormente al chiasma ottico. Ciascun nucleo unilaterale contiene circa 10.000 neuroni divisi in un core ventrale (che supera il chiasma e riceve informazioni dalla retina) e una shell dorsale (che si avvolge parzialmente e riceve gli input dal core). L’SCN è composto anche da cellule gliali che possono potenzialmente contribuire alle funzioni pacemaker. Infatti gli astrociti hanno mostrato dei ritmi circadiani di espressione genica.
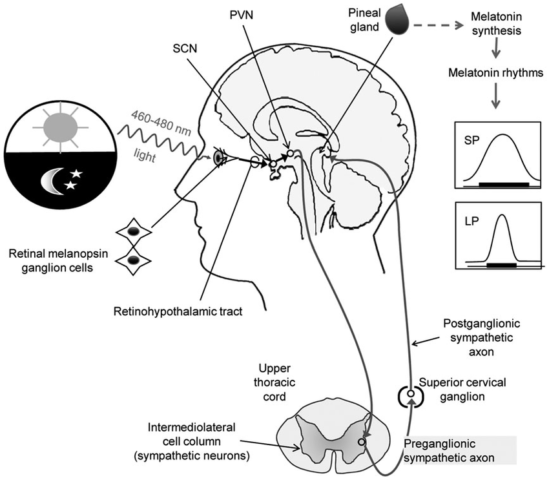
Dalla retina all’epifisi: regolazione della melatonina
La luce influenza la secrezione di melatonina da parte della ghiandola pineale grazie a un circuito neuronale multisinaptico indiretto. La luce penetra nell’occhio mediante il foro pupillare, stimola la retina e invia i segnali al pacemaker circadiano (il nucleo soprachiasmatico ipotalamico) attraverso delle proiezioni retinoipotalamiche dirette (RHT). Un secondo sistema neurale in grado di influenzare l’attività di SCN in base al quantitativo di luce fa affidamento sul foglietto intergenicolato del talamo (nucleo nel complesso talamico laterale circondato dorsalmente per quasi tutta la sua lunghezza dal nucleo genicolato laterale dorsale e ventralmente dal nucleo genicolato laterale ventrale). Dal SCN, gli impulsi viaggiano verso la ghiandola pineale, contraendo prima sinapsi a livello dei nuclei paraventricolari dell’ipotalamo, discendendo poi per sinaptare nella colonna intermediolaterale del tratto spinale toracico superiore, riproiettandosi al ganglio cervicale superiore, e risalendo accanto ai vasi cerebrali principali fino a raggiungere la ghiandola pineale.
Specificamente, i nervi che lasciano il ganglio cervicale precedentemente descritto, risalgono vicino alla carotide interna. Gli assoni simpatici raggiungono la superficie della ghiandola pineale mediante i nervi conari, che sono dettagliatamente delle fibre amieliniche noradrenergiche passanti per il tentorio del cervelletto e aventi nuclei nel ganglio cervicale superiore.